


Unexpected Wound Effects of Local Arginine Infusion:
Reduced Granulation Tissue Formation and Elevated Homocysteine Concentrations
Abstract: Background. Therapeutic use of supplemental arginine has been proposed as an efficacious method to produce nitric oxide (NO) from nitric oxide synthase (NOS) and proline and polyamines from arginase to improve wound healing. This study was designed to examine the effects of arginine on wound angiogenesis and granulation tissue formation. Methods. A ventral hernia, surgically created in the abdominal wall of 12 swine, was repaired with silicone sheeting and skin closure. An osmotic infusion pump, inserted in a remote subcutaneous pocket, continuously delivered saline solution (n = 6) or L-arginine (n = 6) into the wound environment. Granulation tissue thickness was determined by ultrasonography. Fluid was aspirated serially from the developing wound compartment for measurement of nitrite/nitrate (NOx) and amino acid concentrations. On day 14, the animals were sacrificed, and the abdominal wall was harvested for histologic analysis. Results. In animals that received saline, a 4-fold increase in granulation tissue thickness was measured during the 14-day interval. In contrast, in L-arginine treated animals, the day 14 granulation tissue thickness was unchanged from day 4 values of saline treated animals (10.1 mm ± 1.1 mm versus 20.2 mm ± 1.7 mm at day 14; P 0.05). Wound vessel count and vascular surface area estimates derived from image analysis of histologic sections were 2- to 3-fold lower in L-arginine animals compared to controls (P 0.05). Progressive and sustained increases in wound fluid NOx and homocysteine levels were noted in L-arginine treated animals compared to controls (230 µm/L versus 75 µm/L at day 14 [P 0.05]; peak 25.2 µm/L versus 17.3 µm/L at day 7 [P 0.05], respectively). Conclusion. Supplemental arginine induces sustained NO production and creates a methylation demand, resulting in elevated homocysteine concentrations with consequent reductions in wound angiogenesis and granulation tissue formation.
Address correspondence to: Thomas R. Howdieshell, MD Trauma/Surgical Critical Care Department of Surgery University of New Mexico HSC MSC10-5610 Albuquerque, NM 87131 Phone: 505-272-6441 E-mail: thowdieshell@salud.unm.edu
In addition to its role in protein synthesis and nitrogen disposal, arginine serves as a precursor to glutamine, proline, and putrescine (via ornithine), with the latter compound participating in the synthesis of polyamines.1 However, the functional role that has attracted the greatest interest to physicians caring for injured and critically ill patients is the contribution of arginine in the synthesis of nitric oxide (NO). Three distinct human nitric oxide synthase (NOS) enzymes (neuronal NOS or NOS 1, inducible NOS or NOS 2, and endothelial NOS or NOS 3) have been identified, with differing cellular distribution and regulatory mechanisms. All NOS enzymes use L-arginine and molecular oxygen as substrates for the production of NO and L-citrulline. NOS 1 and NOS 3 are constitutively regulated, whereas NOS 2 is transcriptionally regulated by cytokines, endotoxin, and hypoxia. 2 Nitric oxide is a critical mediator of normal tissue repair. Angiogenesis, granulation tissue formation, epidermal migration, and collagen deposition are all significant wound repair processes that are regulated by NO bioactivity. 3–6 The authors have reported previously that NOS 2 plays a critical role in wound granulation tissue formation, and provided in-vivo evidence for dramatic reductions in wound fluid and tissue vascular endothelial growth factor (VEGF) protein content during healing in pigs treated with a selective NOS 2 inhibitor. 7,8 These data suggested an important role for NO in regulating growth factor mediated processes during wound repair, and therefore, identified NO as a potential therapeutic target molecule to improve disorders of wound healing. Repair of excisional wounds is impaired in conditions associated with reduced NOS 2 expression and NO availability, such as steroid administration and diabetes mellitus. 9,10 Therefore, it was hypothesized that supplemental arginine might increase wound levels of NO with resultant increases in granulation tissue formation. Effects of L-Arginine on wound NOx production and amino acid content. Nitrite and nitrate (NOx) are the oxidation products of nitric oxide (NO) in biologic fluids and are used as an index of NOS activity. 15 In saline-treated animals, there was a significant increase in the production of NOx in wound fluid, which peaked between days 7 and 11, and decreased almost to baseline levels by day 14. A significant and sustained increase in wound fluid NOx production was noted in L-arginine treated animals compared to saline controls (230 µM versus 75 µM at day 14; P 0.05, [Figure 2]).
Serial quantitation of amino acid content in wound fluid revealed reductions in L-arginine concentrations and increases in L-ornithine concentrations during the 14-day study interval. However, over the entire time course, L-arginine concentrations remained 2- to 4-fold greater in L-arginine treated animals compared to controls (P 0.05); there were no significant differences in L-ornithine concentrations noted between groups (Figures 3A, B). L-citrulline, L-proline, and L-hydroxyproline levels remained essentially unchanged over time, with no differences noted between groups (data not shown).
Effects of L-Arginine on wound NOx production and amino acid content. Nitrite and nitrate (NOx) are the oxidation products of nitric oxide (NO) in biologic fluids and are used as an index of NOS activity. 15 In saline-treated animals, there was a significant increase in the production of NOx in wound fluid, which peaked between days 7 and 11, and decreased almost to baseline levels by day 14. A significant and sustained increase in wound fluid NOx production was noted in L-arginine treated animals compared to saline controls (230 µM versus 75 µM at day 14; P 0.05, [Figure 2]).
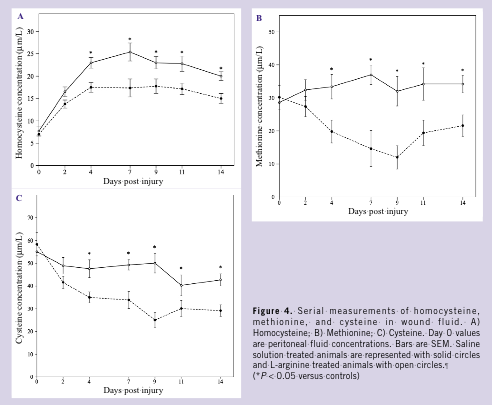
Serial quantitation of amino acid content in wound fluid revealed reductions in L-arginine concentrations and increases in L-ornithine concentrations during the 14-day study interval. However, over the entire time course, L-arginine concentrations remained 2- to 4-fold greater in L-arginine treated animals compared to controls (P 0.05); there were no significant differences in L-ornithine concentrations noted between groups (Figures 3A, B). L-citrulline, L-proline, and L-hydroxyproline levels remained essentially unchanged over time, with no differences noted between groups (data not shown).  Progressive increases in levels of homocysteine were noted in control and L-arginine wound fluids (Figure 4A). However, homocysteine concentrations were significantly higher in L-arginine treated animals compared to saline controls (peak, 25.2 µM/L versus 17.3 µM/L at day 7; P 0.05). Methionine and cysteine concentrations (Figures 4B, C), molecules important in homocysteine metabolism, were significantly higher in L-arginine treated animals compared to controls (peak, 38 µM/L versus 16 µM/L at day 7; P 0.05; 50 µM/L versus 25 µM/L at day 9; P 0.05, respectively). Control and treatment group blood and peritoneal fluid levels of homocysteine, methionine, and cysteine were nearly identical, and therefore, day 0 peritoneal fluid values are reported as controls.
Progressive increases in levels of homocysteine were noted in control and L-arginine wound fluids (Figure 4A). However, homocysteine concentrations were significantly higher in L-arginine treated animals compared to saline controls (peak, 25.2 µM/L versus 17.3 µM/L at day 7; P 0.05). Methionine and cysteine concentrations (Figures 4B, C), molecules important in homocysteine metabolism, were significantly higher in L-arginine treated animals compared to controls (peak, 38 µM/L versus 16 µM/L at day 7; P 0.05; 50 µM/L versus 25 µM/L at day 9; P 0.05, respectively). Control and treatment group blood and peritoneal fluid levels of homocysteine, methionine, and cysteine were nearly identical, and therefore, day 0 peritoneal fluid values are reported as controls.
 Reductions in granulation tissue growth and vascularity. The day 14 granulation tissue harvested from L-arginine infused wounds was remarkably thin, friable, and uniformly white in color compared to the saline infused wounds. All skin incisions healed without evidence of wound infection. Quantitative ultrasound scan analysis detected significant reductions in L-arginine infused granulation tissue thickness compared with controls between days 4 and 14 (Figure 1). Morphometric measurement of day 0 soft tissue and day 14 granulation tissue in L-arginine and control animals confirmed the accuracy of ultrasonography in determination of granulation tissue thickness.
Histologic analysis revealed that the tissue was composed primarily of inflammatory cells and was strikingly devoid of vasculature. Counts from sections immunostained for von Willebrand’s factor, in which endothelial cells could be unambiguously identified, confirmed that L-arginine treatment resulted in 2- to 3-fold reductions in granulation tissue vessel count (control, 2850 ± 310; L-arginine, 1210 ± 250; P 0.05) and luminal cross-sectional surface area (control, 1.28 mm2 ± 0.22 mm2; L-arginine, 0.41 mm2 ± 0.10 mm2; P 0.05) compared with control wounds. Histologic examination of granulation vasculature revealed identical morphology of vascular anatomy in L-arginine and saline treated animals, confirming that the reduced vascular counts in the L-arginine animals were due to fewer vessels, not less tortuous vasculature.
Reductions in granulation tissue growth and vascularity. The day 14 granulation tissue harvested from L-arginine infused wounds was remarkably thin, friable, and uniformly white in color compared to the saline infused wounds. All skin incisions healed without evidence of wound infection. Quantitative ultrasound scan analysis detected significant reductions in L-arginine infused granulation tissue thickness compared with controls between days 4 and 14 (Figure 1). Morphometric measurement of day 0 soft tissue and day 14 granulation tissue in L-arginine and control animals confirmed the accuracy of ultrasonography in determination of granulation tissue thickness.
Histologic analysis revealed that the tissue was composed primarily of inflammatory cells and was strikingly devoid of vasculature. Counts from sections immunostained for von Willebrand’s factor, in which endothelial cells could be unambiguously identified, confirmed that L-arginine treatment resulted in 2- to 3-fold reductions in granulation tissue vessel count (control, 2850 ± 310; L-arginine, 1210 ± 250; P 0.05) and luminal cross-sectional surface area (control, 1.28 mm2 ± 0.22 mm2; L-arginine, 0.41 mm2 ± 0.10 mm2; P 0.05) compared with control wounds. Histologic examination of granulation vasculature revealed identical morphology of vascular anatomy in L-arginine and saline treated animals, confirming that the reduced vascular counts in the L-arginine animals were due to fewer vessels, not less tortuous vasculature.
 In addition to its role as a physiological mediator of numerous cellular and organ functions, NO at high concentrations inhibits cell proliferation. 17 NO inhibits two critical enzymes in the arginine-polyamine pathway. Arginase, which catalyzes the conversion of arginine to ornithine and urea, is important not only in the urea cycle in the liver, but also in biochemical pathways essential to cell growth in all cells. 18 Ornithine is, in turn, converted to putrescine by the enzyme ornithine decarboxylase (ODC), after which putrescine is converted to spermidine and spermine. These 3 polyamines are required for mammalian cell growth. 19 NO has been found to be an inhibitor of ODC, thereby limiting polyamine synthesis. 20 N-hydroxyarginine (NOHA), the principal intermediate in the NOS-catalyzed conversion of arginine to NO and citrulline, is a known competitive inhibitor of arginase and limits ornithine production. 21
Therefore, sustained NO and NOHA production might be inhibiting ODC and arginase, thereby limiting polyamine and ornithine synthesis. Reduced polyamine levels could be responsible for decreased fibroblast and endothelial cell proliferation, which are key cellular constituents of granulation tissue. Reduced ornithine levels, an important substrate not only in polyamine production but also in proline synthesis, a required element of collagen, could be responsible for decreased matrix composition of granulation tissue.
Creatine synthesis is another well known pathway for arginine catabolism. 22 The guanidine group of arginine is transferred to glycine to form guanidinoacetate (GAA) and ornithine by the enzyme arginine: glycine amidinotransferase (AGAT), principally localized to the kidney and pancreas. 23 In turn, GAA is taken up by the liver where it undergoes methylation to creatine via S-adenosyl-L-methionine: GAA N-methyltransferase (GAMT), which converts S-adenosylmethionine into S-adenosylhomocysteine (SAH) in the process. S-adenosylhomocysteine is reversibly hydrolyzed into homocysteine and adenosine. Therefore, the genesis of creatine from arginine creates a methylation demand (Figure 5A) and resultant increase in homocysteine production. 24
Evaluated serum homocysteine may inhibit acute wound repair by altering normal thrombosis and by binding to fibronectin. In normal individuals, greater than 70% of circulating homocysteine is disulfide-bonded to plasma proteins. 25 Fibronectin, a multifunctional glycoprotein important in adhesion, migration, embryogenesis, hemostasis, and wound healing, has free cysteine residues and numerous disulfide bonds. 26 Homocysteine binds to fibronectin resulting in a functional change—inhibition of fibrin binding. 27
The binding of fibronectin to fibrin is vital in thrombosis and wound healing. Tissue injury produces a rapid induction of the clotting cascade and the formation of a provisional matrix, the major components of which are fibrin and fibronectin. 28 The provisional matrix serves as a substrate for the adhesion and migration of mesenchymal cells. Fibroblasts and endothelial cells migrate from the wound area to the provisional matrix as the rate-limiting step in granulation tissue formation. 29 Therefore, a functional change in fibronectin may result in impaired granulation.
Endothelial dysfunction refers to impairment of the normal hemostatic and proliferative properties of vascular endothelium including regulation of vascular tone, hemostasis, inflammation, and angiogenesis. 30 Reported mechanisms by which elevated homocysteine induces endothelial dysfunction include an increase in the procoagulant activity of endothelial cells due to increased tissue factor activity, inhibition of protein C activation, aberrant processing and secretion of thrombomodulin, and reduction in cellular binding sites for tissue plasminogen activator. 31–34
In addition to its prothrombotic effects, homocysteine alters the vasoactive properties of endothelial cells by impairing the production of NO and by decreasing NO bioavailability through alternative mechanisms, such as accelerated oxidative inactivation of NO and “uncoupling” of NOS activity. 35,36 Finally, recent reports suggest that clinically relevant concentrations of homocysteine (10–40 µM/L) inhibit endothelial cell growth by transcriptional inhibition of the cyclin A gene; a gene that is important in cell cycle transitions. 37 Any of these mechanisms may lead to impaired wound angiogenesis.
Finally, homocysteine is normally remethylated to methionine and eliminated by transsulfuration to cysteine (Figure 5B). As endothelial cells are not capable of transsulfuration, the resulting increase in local homocysteine concentration might contribute to wound endothelial dysfunction and injury, and ultimately to impaired granulation tissue formation. 38 Recently, vitamin therapy utilizing folate, B6, and B12, cofactors that are essential in the remethylation and transsulfuration pathways, reduced elevated serum homocysteine concentrations, improved NO bioavailability, accelerated healing of chronic diabetic lower extremity wounds, and improved diabetic peripheral neuropathy. 39–41 Therefore, homocysteine, a known independent risk factor for atherothrombosis, 42 may be an important risk factor for impaired wound healing. The authors are planning further investigation into its production, catabolism, and activity in the wound environment.
From the Department of Surgery, Health Sciences Center, University of New Mexico, Albuquerque
Disclosure: Supported by American Heart Association Southeast Affiliate (T.R.H.)
In addition to its role as a physiological mediator of numerous cellular and organ functions, NO at high concentrations inhibits cell proliferation. 17 NO inhibits two critical enzymes in the arginine-polyamine pathway. Arginase, which catalyzes the conversion of arginine to ornithine and urea, is important not only in the urea cycle in the liver, but also in biochemical pathways essential to cell growth in all cells. 18 Ornithine is, in turn, converted to putrescine by the enzyme ornithine decarboxylase (ODC), after which putrescine is converted to spermidine and spermine. These 3 polyamines are required for mammalian cell growth. 19 NO has been found to be an inhibitor of ODC, thereby limiting polyamine synthesis. 20 N-hydroxyarginine (NOHA), the principal intermediate in the NOS-catalyzed conversion of arginine to NO and citrulline, is a known competitive inhibitor of arginase and limits ornithine production. 21
Therefore, sustained NO and NOHA production might be inhibiting ODC and arginase, thereby limiting polyamine and ornithine synthesis. Reduced polyamine levels could be responsible for decreased fibroblast and endothelial cell proliferation, which are key cellular constituents of granulation tissue. Reduced ornithine levels, an important substrate not only in polyamine production but also in proline synthesis, a required element of collagen, could be responsible for decreased matrix composition of granulation tissue.
Creatine synthesis is another well known pathway for arginine catabolism. 22 The guanidine group of arginine is transferred to glycine to form guanidinoacetate (GAA) and ornithine by the enzyme arginine: glycine amidinotransferase (AGAT), principally localized to the kidney and pancreas. 23 In turn, GAA is taken up by the liver where it undergoes methylation to creatine via S-adenosyl-L-methionine: GAA N-methyltransferase (GAMT), which converts S-adenosylmethionine into S-adenosylhomocysteine (SAH) in the process. S-adenosylhomocysteine is reversibly hydrolyzed into homocysteine and adenosine. Therefore, the genesis of creatine from arginine creates a methylation demand (Figure 5A) and resultant increase in homocysteine production. 24
Evaluated serum homocysteine may inhibit acute wound repair by altering normal thrombosis and by binding to fibronectin. In normal individuals, greater than 70% of circulating homocysteine is disulfide-bonded to plasma proteins. 25 Fibronectin, a multifunctional glycoprotein important in adhesion, migration, embryogenesis, hemostasis, and wound healing, has free cysteine residues and numerous disulfide bonds. 26 Homocysteine binds to fibronectin resulting in a functional change—inhibition of fibrin binding. 27
The binding of fibronectin to fibrin is vital in thrombosis and wound healing. Tissue injury produces a rapid induction of the clotting cascade and the formation of a provisional matrix, the major components of which are fibrin and fibronectin. 28 The provisional matrix serves as a substrate for the adhesion and migration of mesenchymal cells. Fibroblasts and endothelial cells migrate from the wound area to the provisional matrix as the rate-limiting step in granulation tissue formation. 29 Therefore, a functional change in fibronectin may result in impaired granulation.
Endothelial dysfunction refers to impairment of the normal hemostatic and proliferative properties of vascular endothelium including regulation of vascular tone, hemostasis, inflammation, and angiogenesis. 30 Reported mechanisms by which elevated homocysteine induces endothelial dysfunction include an increase in the procoagulant activity of endothelial cells due to increased tissue factor activity, inhibition of protein C activation, aberrant processing and secretion of thrombomodulin, and reduction in cellular binding sites for tissue plasminogen activator. 31–34
In addition to its prothrombotic effects, homocysteine alters the vasoactive properties of endothelial cells by impairing the production of NO and by decreasing NO bioavailability through alternative mechanisms, such as accelerated oxidative inactivation of NO and “uncoupling” of NOS activity. 35,36 Finally, recent reports suggest that clinically relevant concentrations of homocysteine (10–40 µM/L) inhibit endothelial cell growth by transcriptional inhibition of the cyclin A gene; a gene that is important in cell cycle transitions. 37 Any of these mechanisms may lead to impaired wound angiogenesis.
Finally, homocysteine is normally remethylated to methionine and eliminated by transsulfuration to cysteine (Figure 5B). As endothelial cells are not capable of transsulfuration, the resulting increase in local homocysteine concentration might contribute to wound endothelial dysfunction and injury, and ultimately to impaired granulation tissue formation. 38 Recently, vitamin therapy utilizing folate, B6, and B12, cofactors that are essential in the remethylation and transsulfuration pathways, reduced elevated serum homocysteine concentrations, improved NO bioavailability, accelerated healing of chronic diabetic lower extremity wounds, and improved diabetic peripheral neuropathy. 39–41 Therefore, homocysteine, a known independent risk factor for atherothrombosis, 42 may be an important risk factor for impaired wound healing. The authors are planning further investigation into its production, catabolism, and activity in the wound environment.
From the Department of Surgery, Health Sciences Center, University of New Mexico, Albuquerque
Disclosure: Supported by American Heart Association Southeast Affiliate (T.R.H.)
Address correspondence to: Thomas R. Howdieshell, MD Trauma/Surgical Critical Care Department of Surgery University of New Mexico HSC MSC10-5610 Albuquerque, NM 87131 Phone: 505-272-6441 E-mail: thowdieshell@salud.unm.edu
In addition to its role in protein synthesis and nitrogen disposal, arginine serves as a precursor to glutamine, proline, and putrescine (via ornithine), with the latter compound participating in the synthesis of polyamines.1 However, the functional role that has attracted the greatest interest to physicians caring for injured and critically ill patients is the contribution of arginine in the synthesis of nitric oxide (NO). Three distinct human nitric oxide synthase (NOS) enzymes (neuronal NOS or NOS 1, inducible NOS or NOS 2, and endothelial NOS or NOS 3) have been identified, with differing cellular distribution and regulatory mechanisms. All NOS enzymes use L-arginine and molecular oxygen as substrates for the production of NO and L-citrulline. NOS 1 and NOS 3 are constitutively regulated, whereas NOS 2 is transcriptionally regulated by cytokines, endotoxin, and hypoxia. 2 Nitric oxide is a critical mediator of normal tissue repair. Angiogenesis, granulation tissue formation, epidermal migration, and collagen deposition are all significant wound repair processes that are regulated by NO bioactivity. 3–6 The authors have reported previously that NOS 2 plays a critical role in wound granulation tissue formation, and provided in-vivo evidence for dramatic reductions in wound fluid and tissue vascular endothelial growth factor (VEGF) protein content during healing in pigs treated with a selective NOS 2 inhibitor. 7,8 These data suggested an important role for NO in regulating growth factor mediated processes during wound repair, and therefore, identified NO as a potential therapeutic target molecule to improve disorders of wound healing. Repair of excisional wounds is impaired in conditions associated with reduced NOS 2 expression and NO availability, such as steroid administration and diabetes mellitus. 9,10 Therefore, it was hypothesized that supplemental arginine might increase wound levels of NO with resultant increases in granulation tissue formation.
Methods
Surgical procedures. All animals were treated humanely in accordance with the National Research Council’s Guide for the Care and Use of Laboratory Animals as part of a protocol approved by the University of New Mexico’s animal review committee. Domestic female Landrace swine (n = 12; weight = 15 kg–20 kg) underwent induction with Telazol, xylazine hydrochloride, and atropine. The animals were anesthetized with isoflurane (1.5% to 3%) via inhalation through an endotracheal tube. Postsurgical analgesia was provided using buprenorphine hydrochloride for 48 hours. After maintenance of inhalation anesthesia, sterile preparation (povidone-iodine), and draping, an infraumbilical diagnostic peritoneal lavage was performed (Arrow DPL Kit, Arrow International, Reading, PA) with the percutaneous Seldinger technique to obtain preincision peritoneal fluid for biochemical and molecular analysis. A sample of plasma was obtained from an ear vein. Next, a supraumbilical midline laparotomy was performed, and biopsy samples of omentum, peritoneum, and portions of abdominal wall were taken for use as controls. A full-thickness section of the abdominal wall (8 cm x 12 cm), excluding the skin, was surgically excised to create a hernia defect. A similar-sized piece of silicone sheeting (BioPlexus Corp, Saticoy, CA) was sutured to the fascial edges with monofilament suture to effect abdominal wall closure. Two osmotic minipumps with attached polyethylene tubing were surgically implanted into each pig into remote abdominal wall subcutaneous pockets (model ML-2; Alzet Corp, Palo Alto, CA). The attached tubing, used to deliver saline solution (n = 6) or L-arginine (n = 6) into the wound environment, entered the wound compartment via a subcutaneous tunnel. To ensure uniform delivery of L-arginine or saline solution within the aqueous wound compartment, the tubing contained multiple side holes and was looped to increase surface area. The pump was manufactured to deliver 5 µL/h for 14 days. Before implantation, the pump was loaded with saline solution or L-arginine under sterile conditions and maintained at 37˚C in an incubator to ensure optimal delivery. After pump insertion, the midline and pocket skin incisions were closed in 2 layers to cover the biomaterial and pumps, and thus, complete wound closure. Each animal was housed for 14 days after surgery and received water and food ad libitum. On days 2, 4, 7, 9, 11, and 14 after surgery, each animal underwent sedation, was placed supine, and abdominal ultrasonography was performed with an Acusom Ultrasound Imager with 7.5-mHz linear array and 3.5-mHz sector probe (Acusom Corp, Mountain View, CA). At each time point, images that were recorded electronically for later analysis included granulation tissue thickness (measured in millimeters) with cursors marking multiple consistent sites. Each day, a sample of wound fluid located in the compartment between the skin and silicone sheeting was percutaneously aspirated with a 23-gauge needle under sterile conditions, and guided by an ultrasound scan. On day 14 after surgery, the animals underwent sedation, and an infraumbilical diagnostic peritoneal lavage was performed to obtain presacrifice peritoneal fluid for assay. A blood sample was obtained from an ear vein. Then each animal was sacrificed with pentobarbital, and the abdominal wall was harvested en bloc. The osmotic minipumps were removed from the subcutaneous pockets, and their residual volume was measured to confirm complete delivery. Histologic and biochemical analyses were performed on blood, peritoneal fluid, local wound fluid, and subcutaneous granulation tissue. Nitrite/nitrate (NOx) measurement. NOx was assayed in wound and peritoneal fluids with the Bioxytech NO nonenzymatic assay (Oxis International Inc, Portland, OR) according to the manufacturer’s instructions. All fluids were centrifuged immediately after harvest and frozen at -70˚C until they were assayed. Each sample was run in duplicate and at pertinent dilutions, with mean values reported. Quantitation by high-pressure liquid chromatography. Amino acids (arginine, methionine, cysteine, ornithine, proline, hydroxyproline, and citrulline) were quantified with high-pressure liquid chromatography (DX500, Dionex Inc, Sunnyvale, CA) using a Pickering post-column ninhydrin reactor (Pickering Corp, Mountain View, CA). Homocysteine quantitation. Homocysteine was quantified with a solid-phase, competing chemiluminescent enzyme immunoassay according to the manufacturer’s instructions (Immulite 2500, Siemens Medical Solutions Diagnostics, Deerfield, IL). Briefly, peritoneal or wound fluid samples were pretreated with S-adenosyl-L-homocysteine (SAH) hydrolase and dithiothreitol (DTT) solution in a reaction tube containing no bead. After a 30-minute incubation, the treated sample was transferred to a second reaction tube containing a SAH-coated polystyrene bead and an alkaline phosphatase-labeled antibody specific for SAH. During a 30-minute incubation, the converted SAH from the sample pretreatment competed with immobilized SAH for binding alkaline phosphatase-labeled anti-SAH antibody. Unbound enzyme conjugate was removed by centrifugal wash. Substrate was added and the procedure continued as a typical immunoassay. The documented analytical sensitivity of this method is 0.5 µM/L. 11 Vascular density and morphometric determinations. Granulation tissue vessel count and vascular luminal cross-sectional surface area were determined with image analysis of von Willebrand’s factor-immunostained sections. Multiple consistent wound biopsies were analyzed. The boundaries of the granulation tissue region to be analyzed were the wound fluid compartment and distinct subcutaneous tissue interface. Immunohistochemical analysis was performed using the avidin-biotin method, as previously reported. 12 A Zeiss microscope with attached digital camera was used for image analysis. The magnified image (x200) of the slide section was acquired with Adobe Photoshop and analyzed with IPLab Spectrum software on an Apple Macintosh computer. Vessels in each section were defined by the circular or ovoid image of the brown endothelial walls. Capillaries, arterioles, and venules were counted. Vessels were identified and marked by an observer with no knowledge of treatment group. The software program was used to count the number of vessels marked in each section. The cross-sectional luminal area of vessels was computed to confirm the vascular density estimates. Closed vessels would alter the numeric count, but only minimally affect the estimate of vascular area. Thus, the determination of luminal area constituted a separate estimate of vascular density. The minimum and maximum major axis of each vessel was measured to determine the area. The calibration scale was set with stage micrometer to match computer pixels to a micrometer scale. The cross-sectional area of each measurement field was computed with careful outlining of the previously reported boundaries of granulation tissue. 12 Granulation tissue thickness was also determined with morphometric analysis of histologic specimens for confirmation of ultrasonographic measurements. The wound biopsies used for vascular density determination were scanned with the previously described image analysis techniques. Granulation tissue thickness was measured in millimeters by cursor-defined boundaries with an established calibration scale and software computations. L-arginine supplementation. L-arginine was purchased from Alexis Biochemicals (San Diego, CA). Each osmotic infusion pump (2 per pig) was loaded with either 2-mL of saline solution (n = 6) or a 2-mL solution containing 50 mg/mL of L-arginine in saline solution as a carrier (n = 6). Each pump was manufactured to deliver 5 µL/h for 14 days. The L-arginine dose was based on maximal solubility of arginine in physiologic saline.Statistical Analysis
All data are expressed as mean ± the standard error of the mean (SEM). Differences among groups and between baseline and subsequent time points were determined with repeated-measures analysis of variance with Tukey significant difference test used for post hoc analysis; P ≤ 0.05 was considered statistically significant.Results
Granulation tissue growth, angiogenesis, and wound fluid accumulation. To provide a readily quantitated wound healing response, a large segment of abdominal wall, including fascia, muscle, and peritoneum (excluding skin) was surgically excised from pigs and replaced with silicone sheeting. The sheeting does not incorporate into adjacent tissues, but rather becomes encapsulated and creates an excellent scaffold for granulation tissue formation and local wound fluid accumulation. 13,14 Delivery of saline solution into the wound compartment did not alter the predicted wound responses. 7,13 The skin incision was completely epithelialized by day 14, without evidence of induration or erythema. The subcutaneous granulation tissue harvested at day 14 from animals that received saline solution was thick, tough, and red in appearance. Sonography was used to serially image the wound site to measure the thickness of the developing granulation tissue. A nearly linear increase in granulation tissue thickness was measured from serially acquired images, and by day 14, the granulation tissue had reached a thickness of 20.2 mm ± 1.2 mm in saline treated animals (Figure 1). Histologically, control group granulation tissue (day 14) was observed to consist of a neutrophil/macrophage-rich zone that bordered on the wound fluid compartment, a microvascular-rich zone whose endothelial cell components could be stained with an antibody against von Willebrand’s factor, and a fibroblast-rich, most mature zone.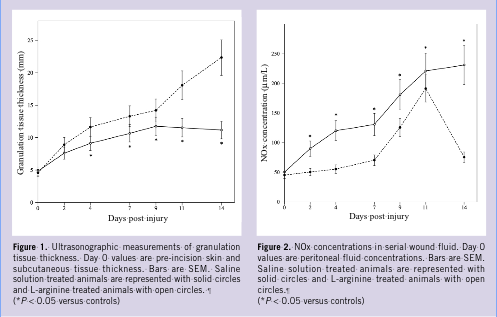 Effects of L-Arginine on wound NOx production and amino acid content. Nitrite and nitrate (NOx) are the oxidation products of nitric oxide (NO) in biologic fluids and are used as an index of NOS activity. 15 In saline-treated animals, there was a significant increase in the production of NOx in wound fluid, which peaked between days 7 and 11, and decreased almost to baseline levels by day 14. A significant and sustained increase in wound fluid NOx production was noted in L-arginine treated animals compared to saline controls (230 µM versus 75 µM at day 14; P 0.05, [Figure 2]).
Serial quantitation of amino acid content in wound fluid revealed reductions in L-arginine concentrations and increases in L-ornithine concentrations during the 14-day study interval. However, over the entire time course, L-arginine concentrations remained 2- to 4-fold greater in L-arginine treated animals compared to controls (P 0.05); there were no significant differences in L-ornithine concentrations noted between groups (Figures 3A, B). L-citrulline, L-proline, and L-hydroxyproline levels remained essentially unchanged over time, with no differences noted between groups (data not shown).
Effects of L-Arginine on wound NOx production and amino acid content. Nitrite and nitrate (NOx) are the oxidation products of nitric oxide (NO) in biologic fluids and are used as an index of NOS activity. 15 In saline-treated animals, there was a significant increase in the production of NOx in wound fluid, which peaked between days 7 and 11, and decreased almost to baseline levels by day 14. A significant and sustained increase in wound fluid NOx production was noted in L-arginine treated animals compared to saline controls (230 µM versus 75 µM at day 14; P 0.05, [Figure 2]).
Serial quantitation of amino acid content in wound fluid revealed reductions in L-arginine concentrations and increases in L-ornithine concentrations during the 14-day study interval. However, over the entire time course, L-arginine concentrations remained 2- to 4-fold greater in L-arginine treated animals compared to controls (P 0.05); there were no significant differences in L-ornithine concentrations noted between groups (Figures 3A, B). L-citrulline, L-proline, and L-hydroxyproline levels remained essentially unchanged over time, with no differences noted between groups (data not shown). 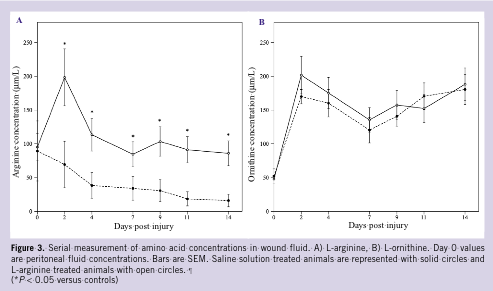 Progressive increases in levels of homocysteine were noted in control and L-arginine wound fluids (Figure 4A). However, homocysteine concentrations were significantly higher in L-arginine treated animals compared to saline controls (peak, 25.2 µM/L versus 17.3 µM/L at day 7; P 0.05). Methionine and cysteine concentrations (Figures 4B, C), molecules important in homocysteine metabolism, were significantly higher in L-arginine treated animals compared to controls (peak, 38 µM/L versus 16 µM/L at day 7; P 0.05; 50 µM/L versus 25 µM/L at day 9; P 0.05, respectively). Control and treatment group blood and peritoneal fluid levels of homocysteine, methionine, and cysteine were nearly identical, and therefore, day 0 peritoneal fluid values are reported as controls.
Progressive increases in levels of homocysteine were noted in control and L-arginine wound fluids (Figure 4A). However, homocysteine concentrations were significantly higher in L-arginine treated animals compared to saline controls (peak, 25.2 µM/L versus 17.3 µM/L at day 7; P 0.05). Methionine and cysteine concentrations (Figures 4B, C), molecules important in homocysteine metabolism, were significantly higher in L-arginine treated animals compared to controls (peak, 38 µM/L versus 16 µM/L at day 7; P 0.05; 50 µM/L versus 25 µM/L at day 9; P 0.05, respectively). Control and treatment group blood and peritoneal fluid levels of homocysteine, methionine, and cysteine were nearly identical, and therefore, day 0 peritoneal fluid values are reported as controls.
 Reductions in granulation tissue growth and vascularity. The day 14 granulation tissue harvested from L-arginine infused wounds was remarkably thin, friable, and uniformly white in color compared to the saline infused wounds. All skin incisions healed without evidence of wound infection. Quantitative ultrasound scan analysis detected significant reductions in L-arginine infused granulation tissue thickness compared with controls between days 4 and 14 (Figure 1). Morphometric measurement of day 0 soft tissue and day 14 granulation tissue in L-arginine and control animals confirmed the accuracy of ultrasonography in determination of granulation tissue thickness.
Histologic analysis revealed that the tissue was composed primarily of inflammatory cells and was strikingly devoid of vasculature. Counts from sections immunostained for von Willebrand’s factor, in which endothelial cells could be unambiguously identified, confirmed that L-arginine treatment resulted in 2- to 3-fold reductions in granulation tissue vessel count (control, 2850 ± 310; L-arginine, 1210 ± 250; P 0.05) and luminal cross-sectional surface area (control, 1.28 mm2 ± 0.22 mm2; L-arginine, 0.41 mm2 ± 0.10 mm2; P 0.05) compared with control wounds. Histologic examination of granulation vasculature revealed identical morphology of vascular anatomy in L-arginine and saline treated animals, confirming that the reduced vascular counts in the L-arginine animals were due to fewer vessels, not less tortuous vasculature.
Reductions in granulation tissue growth and vascularity. The day 14 granulation tissue harvested from L-arginine infused wounds was remarkably thin, friable, and uniformly white in color compared to the saline infused wounds. All skin incisions healed without evidence of wound infection. Quantitative ultrasound scan analysis detected significant reductions in L-arginine infused granulation tissue thickness compared with controls between days 4 and 14 (Figure 1). Morphometric measurement of day 0 soft tissue and day 14 granulation tissue in L-arginine and control animals confirmed the accuracy of ultrasonography in determination of granulation tissue thickness.
Histologic analysis revealed that the tissue was composed primarily of inflammatory cells and was strikingly devoid of vasculature. Counts from sections immunostained for von Willebrand’s factor, in which endothelial cells could be unambiguously identified, confirmed that L-arginine treatment resulted in 2- to 3-fold reductions in granulation tissue vessel count (control, 2850 ± 310; L-arginine, 1210 ± 250; P 0.05) and luminal cross-sectional surface area (control, 1.28 mm2 ± 0.22 mm2; L-arginine, 0.41 mm2 ± 0.10 mm2; P 0.05) compared with control wounds. Histologic examination of granulation vasculature revealed identical morphology of vascular anatomy in L-arginine and saline treated animals, confirming that the reduced vascular counts in the L-arginine animals were due to fewer vessels, not less tortuous vasculature.
Discussion
In this report, osmotic infusion pump delivery of L-arginine into the wound environment and quantitative assessment of its consequences was used to evaluate arginine’s role in wound angiogenesis and granulation tissue formation. Continuous infusion of L-arginine into a local wound environment produced significant and sustained NO production, progressive increases in wound fluid homocysteine levels, and reductions in wound vessel number, vascular surface area, and granulation tissue thickness. Arginine is one of the most versatile amino acids in mammalian cells, serving not only as a precursor for protein synthesis, but also for NO, urea, ornithine, proline, polyamines, and creatine (Figure 5A). Arginine is catabolized to NO and citrulline by action of NOS. Arginine serves as a substrate for arginase, yielding urea and ornithine. Ornithine is an important intermediate in the production of proline, glutamate, and polyamines. 16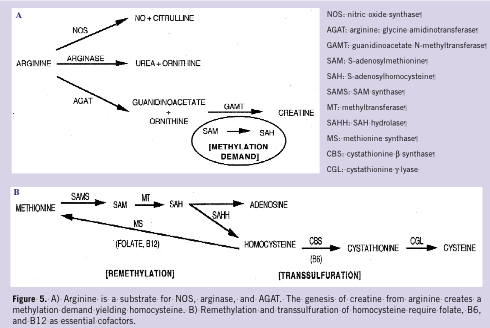 In addition to its role as a physiological mediator of numerous cellular and organ functions, NO at high concentrations inhibits cell proliferation. 17 NO inhibits two critical enzymes in the arginine-polyamine pathway. Arginase, which catalyzes the conversion of arginine to ornithine and urea, is important not only in the urea cycle in the liver, but also in biochemical pathways essential to cell growth in all cells. 18 Ornithine is, in turn, converted to putrescine by the enzyme ornithine decarboxylase (ODC), after which putrescine is converted to spermidine and spermine. These 3 polyamines are required for mammalian cell growth. 19 NO has been found to be an inhibitor of ODC, thereby limiting polyamine synthesis. 20 N-hydroxyarginine (NOHA), the principal intermediate in the NOS-catalyzed conversion of arginine to NO and citrulline, is a known competitive inhibitor of arginase and limits ornithine production. 21
Therefore, sustained NO and NOHA production might be inhibiting ODC and arginase, thereby limiting polyamine and ornithine synthesis. Reduced polyamine levels could be responsible for decreased fibroblast and endothelial cell proliferation, which are key cellular constituents of granulation tissue. Reduced ornithine levels, an important substrate not only in polyamine production but also in proline synthesis, a required element of collagen, could be responsible for decreased matrix composition of granulation tissue.
Creatine synthesis is another well known pathway for arginine catabolism. 22 The guanidine group of arginine is transferred to glycine to form guanidinoacetate (GAA) and ornithine by the enzyme arginine: glycine amidinotransferase (AGAT), principally localized to the kidney and pancreas. 23 In turn, GAA is taken up by the liver where it undergoes methylation to creatine via S-adenosyl-L-methionine: GAA N-methyltransferase (GAMT), which converts S-adenosylmethionine into S-adenosylhomocysteine (SAH) in the process. S-adenosylhomocysteine is reversibly hydrolyzed into homocysteine and adenosine. Therefore, the genesis of creatine from arginine creates a methylation demand (Figure 5A) and resultant increase in homocysteine production. 24
Evaluated serum homocysteine may inhibit acute wound repair by altering normal thrombosis and by binding to fibronectin. In normal individuals, greater than 70% of circulating homocysteine is disulfide-bonded to plasma proteins. 25 Fibronectin, a multifunctional glycoprotein important in adhesion, migration, embryogenesis, hemostasis, and wound healing, has free cysteine residues and numerous disulfide bonds. 26 Homocysteine binds to fibronectin resulting in a functional change—inhibition of fibrin binding. 27
The binding of fibronectin to fibrin is vital in thrombosis and wound healing. Tissue injury produces a rapid induction of the clotting cascade and the formation of a provisional matrix, the major components of which are fibrin and fibronectin. 28 The provisional matrix serves as a substrate for the adhesion and migration of mesenchymal cells. Fibroblasts and endothelial cells migrate from the wound area to the provisional matrix as the rate-limiting step in granulation tissue formation. 29 Therefore, a functional change in fibronectin may result in impaired granulation.
Endothelial dysfunction refers to impairment of the normal hemostatic and proliferative properties of vascular endothelium including regulation of vascular tone, hemostasis, inflammation, and angiogenesis. 30 Reported mechanisms by which elevated homocysteine induces endothelial dysfunction include an increase in the procoagulant activity of endothelial cells due to increased tissue factor activity, inhibition of protein C activation, aberrant processing and secretion of thrombomodulin, and reduction in cellular binding sites for tissue plasminogen activator. 31–34
In addition to its prothrombotic effects, homocysteine alters the vasoactive properties of endothelial cells by impairing the production of NO and by decreasing NO bioavailability through alternative mechanisms, such as accelerated oxidative inactivation of NO and “uncoupling” of NOS activity. 35,36 Finally, recent reports suggest that clinically relevant concentrations of homocysteine (10–40 µM/L) inhibit endothelial cell growth by transcriptional inhibition of the cyclin A gene; a gene that is important in cell cycle transitions. 37 Any of these mechanisms may lead to impaired wound angiogenesis.
Finally, homocysteine is normally remethylated to methionine and eliminated by transsulfuration to cysteine (Figure 5B). As endothelial cells are not capable of transsulfuration, the resulting increase in local homocysteine concentration might contribute to wound endothelial dysfunction and injury, and ultimately to impaired granulation tissue formation. 38 Recently, vitamin therapy utilizing folate, B6, and B12, cofactors that are essential in the remethylation and transsulfuration pathways, reduced elevated serum homocysteine concentrations, improved NO bioavailability, accelerated healing of chronic diabetic lower extremity wounds, and improved diabetic peripheral neuropathy. 39–41 Therefore, homocysteine, a known independent risk factor for atherothrombosis, 42 may be an important risk factor for impaired wound healing. The authors are planning further investigation into its production, catabolism, and activity in the wound environment.
From the Department of Surgery, Health Sciences Center, University of New Mexico, Albuquerque
Disclosure: Supported by American Heart Association Southeast Affiliate (T.R.H.)
In addition to its role as a physiological mediator of numerous cellular and organ functions, NO at high concentrations inhibits cell proliferation. 17 NO inhibits two critical enzymes in the arginine-polyamine pathway. Arginase, which catalyzes the conversion of arginine to ornithine and urea, is important not only in the urea cycle in the liver, but also in biochemical pathways essential to cell growth in all cells. 18 Ornithine is, in turn, converted to putrescine by the enzyme ornithine decarboxylase (ODC), after which putrescine is converted to spermidine and spermine. These 3 polyamines are required for mammalian cell growth. 19 NO has been found to be an inhibitor of ODC, thereby limiting polyamine synthesis. 20 N-hydroxyarginine (NOHA), the principal intermediate in the NOS-catalyzed conversion of arginine to NO and citrulline, is a known competitive inhibitor of arginase and limits ornithine production. 21
Therefore, sustained NO and NOHA production might be inhibiting ODC and arginase, thereby limiting polyamine and ornithine synthesis. Reduced polyamine levels could be responsible for decreased fibroblast and endothelial cell proliferation, which are key cellular constituents of granulation tissue. Reduced ornithine levels, an important substrate not only in polyamine production but also in proline synthesis, a required element of collagen, could be responsible for decreased matrix composition of granulation tissue.
Creatine synthesis is another well known pathway for arginine catabolism. 22 The guanidine group of arginine is transferred to glycine to form guanidinoacetate (GAA) and ornithine by the enzyme arginine: glycine amidinotransferase (AGAT), principally localized to the kidney and pancreas. 23 In turn, GAA is taken up by the liver where it undergoes methylation to creatine via S-adenosyl-L-methionine: GAA N-methyltransferase (GAMT), which converts S-adenosylmethionine into S-adenosylhomocysteine (SAH) in the process. S-adenosylhomocysteine is reversibly hydrolyzed into homocysteine and adenosine. Therefore, the genesis of creatine from arginine creates a methylation demand (Figure 5A) and resultant increase in homocysteine production. 24
Evaluated serum homocysteine may inhibit acute wound repair by altering normal thrombosis and by binding to fibronectin. In normal individuals, greater than 70% of circulating homocysteine is disulfide-bonded to plasma proteins. 25 Fibronectin, a multifunctional glycoprotein important in adhesion, migration, embryogenesis, hemostasis, and wound healing, has free cysteine residues and numerous disulfide bonds. 26 Homocysteine binds to fibronectin resulting in a functional change—inhibition of fibrin binding. 27
The binding of fibronectin to fibrin is vital in thrombosis and wound healing. Tissue injury produces a rapid induction of the clotting cascade and the formation of a provisional matrix, the major components of which are fibrin and fibronectin. 28 The provisional matrix serves as a substrate for the adhesion and migration of mesenchymal cells. Fibroblasts and endothelial cells migrate from the wound area to the provisional matrix as the rate-limiting step in granulation tissue formation. 29 Therefore, a functional change in fibronectin may result in impaired granulation.
Endothelial dysfunction refers to impairment of the normal hemostatic and proliferative properties of vascular endothelium including regulation of vascular tone, hemostasis, inflammation, and angiogenesis. 30 Reported mechanisms by which elevated homocysteine induces endothelial dysfunction include an increase in the procoagulant activity of endothelial cells due to increased tissue factor activity, inhibition of protein C activation, aberrant processing and secretion of thrombomodulin, and reduction in cellular binding sites for tissue plasminogen activator. 31–34
In addition to its prothrombotic effects, homocysteine alters the vasoactive properties of endothelial cells by impairing the production of NO and by decreasing NO bioavailability through alternative mechanisms, such as accelerated oxidative inactivation of NO and “uncoupling” of NOS activity. 35,36 Finally, recent reports suggest that clinically relevant concentrations of homocysteine (10–40 µM/L) inhibit endothelial cell growth by transcriptional inhibition of the cyclin A gene; a gene that is important in cell cycle transitions. 37 Any of these mechanisms may lead to impaired wound angiogenesis.
Finally, homocysteine is normally remethylated to methionine and eliminated by transsulfuration to cysteine (Figure 5B). As endothelial cells are not capable of transsulfuration, the resulting increase in local homocysteine concentration might contribute to wound endothelial dysfunction and injury, and ultimately to impaired granulation tissue formation. 38 Recently, vitamin therapy utilizing folate, B6, and B12, cofactors that are essential in the remethylation and transsulfuration pathways, reduced elevated serum homocysteine concentrations, improved NO bioavailability, accelerated healing of chronic diabetic lower extremity wounds, and improved diabetic peripheral neuropathy. 39–41 Therefore, homocysteine, a known independent risk factor for atherothrombosis, 42 may be an important risk factor for impaired wound healing. The authors are planning further investigation into its production, catabolism, and activity in the wound environment.
From the Department of Surgery, Health Sciences Center, University of New Mexico, Albuquerque
Disclosure: Supported by American Heart Association Southeast Affiliate (T.R.H.)






